|
|
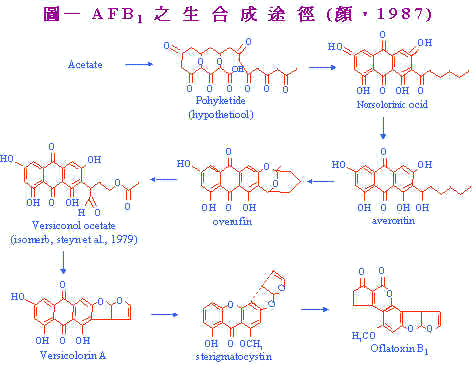
黄麴毒素对鸡只营养分吸收代谢之影响 郑永祥(1) 詹德芳(2) 吴继芳(3) 一、前 言 台湾每年自美国进口大批谷物,其数高达818万公吨(杂粮畜产,1990)谷物自收获,船运、装卸至制成饲料,往往耗时数月,期间之仓贮条件影响谷物之品质;且本省地处热带与亚热带,常年高温多湿,极利於霉菌之繁衍。因此霉菌毒素污染也成为台湾养鸡业及饲料制造业者所高度关注。 黄麴毒素(Aflatoxin)是由Aspergillus flavus及Aspergillus parasiticus所产生的霉菌毒素代谢产物,被认为具有潜在性的肝毒害(hepatoxin)及致癌物(carcinogen)。典型之黄麴毒素中毒症状有嗜眠、垂翼、体温上升和神经症状。造成鸡只生长性能及饲料利用效率降低,胰脏消化酵素活性减低(Osborne et al., 1981),免疫能力减弱(Campbell et al., 1983;Chang et al., 1979),血液凝固延迟(Doerr et al., 1974),肝脏肿大及脂肪移动(mobilized)失调(Huff et al., 1986)等诸多毒害作用对畜禽业者造成莫大的经济损失。 ───────────────────────────────────── (1)国立宜兰农专畜产科 (2)国立中兴大学畜产学系 (3)台湾养猪科学研究所生产系统系 二、黄麴毒素之生合成 黄麴毒素为一族相似,自然存在之鶭喃氧杂鸻邻酮类(furanocoumarins),这些化合物可在食物中黄绿色霉菌Aspergillus fravus及Aspergillus parasiticus之生长而发现,所有之黄麴毒素化合物均是具萤光性;可用长波长紫外线加以观察。 黄麴毒素之生合成;已藉由利用1-C14或2-C14之acetate及methyl-C14 methionine之同位素追踪方法研究其合扇途径;并加以证实。图一即为黄麴毒素B1之生合成径路。利用能阻止AFB1形成之产黄麴毒素变异株及形成其一般代谢性之非产黄麴毒素菌种已可证明此生合成径路中之各中间产物。例如,使用A. parasiticus变异株,得知averantin为AFB1之前驱物质,其位於averufin之前而在norsolorine之後。而versiconal acetate为AFB1之生合成主要中间物,versiconal acetate经酵素作用变成versicolorin A。sterigmatocystin为最後中间产物。 |
|
AFB1及AFG1之构造由Asao於1965年应用X-ray证实其为双dihydrofuran环加以融合成顺式(cis)构形。AFB1合成有不同之方式。AFB2及AFG2可由AFB1及AFG1之dihydrofuran环端还原而成。 Lee et al. (1974)发现在黄麴菌之基础培养其中有锌之存在时;其生长与产毒量均显着增加,如表一。Gupt and Venk itasubramanian (1977)利用放射性元素研究黄麴毒素之合成。发现硫酸锌浓度在10mM时产毒量最高。而植酸(phytic acid)则有抑制作用。此乃为何大豆污染黄麴毒素情形并不严重的原因;因大豆内含有植酸,锌与植酸结合而失去作用所致。Gupta et al. (1976)指出当锌存在时,hexokinase,G-6-P dehydrogenase,aldolase,puruvate kinase及glyceraldehyde-3-phosphate dehydrogenase等糖解酵素活性增高;合成黄麴毒素之前驱物因而增加。 |
| 表一 一 些 无 机 盐 类 对 悬 浮 菌 丝 之 黄 麴 毒 素 比 活 性 影 响 | ||||||
| 添 加 化合物种类 | 黄 麴毒 素 | 添 加 的 浓 度 对 黄 麴 毒 素 比 活 性 改 变 的 % | ||||
| 0.01 | 0.10 | 0.50 | 1.0 | 10.0 | ||
|
硫 酸 锌
硫 酸 锌
氯 化 汞
氯 化 钡
氯 化 钙
氯 化 锰
|
B
G B G B G B G B G B G |
+3
-3 +9 +8 +21 +26 +27 +36 +7 +11 +2 -9 |
+10
0 +15 +14 +17 +17 +27 +37 +9 +15 -8 +13 |
+18
+3 - - -4 +12 +29 +35 +18 +21 +12 +7 |
+17
+4 +17 +15 -2 +9 +34 +30 +21 +27 +7 +9 |
+46
+33 +15 +14 - - +24 +27 +31 +29 +21 +30 |
|
(Gupta et al., 1976) |
||||||
|
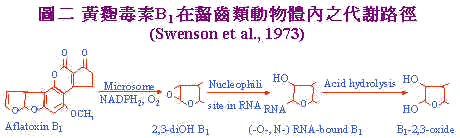
Maggon et al. (1977)指出黄麴菌开始生长时初级代谢即开始进行,此时仅有少量之黄麴毒素:後因磷、氮等初级代谢中所需之养分用罄;生长逐渐受阻,初级代谢物於是累积,如pyruvate,malonate,acetate和各种胺基酸代谢物;引发次级代谢所需之酵素活性,次级代谢作用於焉开始,此时polyketide生合成代谢路径(polyketide biosynthetic pathway)超过脂肪酸的生合成代谢;促进aflatoxin的生合成。 三、黄麴毒素之生体内代谢 黄麴毒素在体内之分解代谢路径可分为径路一和径路二。径路一主要为啮齿类动物(rodents)体内之代谢,如图二 。 |
|
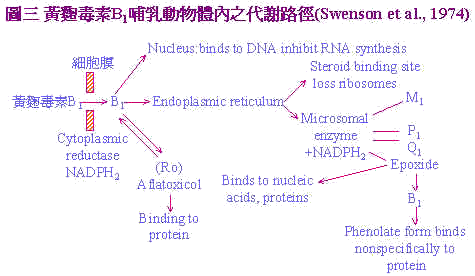
黄麴毒素B1经由饲料进入体内,到达肝脏之微粒体(Microsome)在NADH2及O2存在下氧化为B1-2,3 oxide,此化合物具有高度之亲核性(nucleophilic)会攻击RNA与RNA中之-0-或N-group形成RNA bound B1之产物,再经由酸性水解(Acid hydrolysis)过程转变成毒性较低之2,3-dioHB1。 径路二:主要发生於哺乳类动物:如图3。Aflatoxin B1经由哺乳动物摄入後;通过细胞膜进入细胞,在细胞质还原鶤(cytoplasmic reductase)作用下NADPH2辅助还原为Aflatxicol (Ro)再与蛋白质结合。或进入核仁与DNA键结,继而抑制RNA的合成或经过细胞质中的内质网在微粒体鶤作用及NADPH2存在下代谢成生毒性较弱之AFM1及无毒性AFP1及毒性较强之AFQ1,和最具毒性的Aflatoxin epoxide,此一衍生物可与核酸及蛋白质结合。或epoxide经再代谢成AFB2以石碳酸盐形成与蛋白质呈非特异性的结合。 |
|
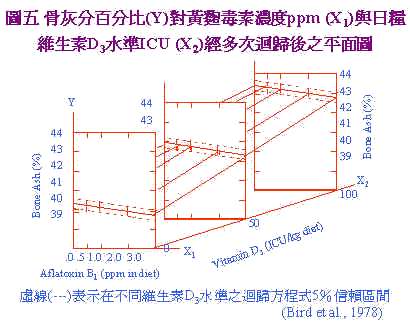
黄麴毒素的影响;对细胞不同层次的作用不一,它对蛋白质形成显然具有抑制作用,因为它与DNA结合阻碍RNA的合成,甚至於所有蛋白质的合成,从转录(transcription)、转译(translation)、直到加长(elongation)各步骤都会发生影响。不过当动物吸收多量及长期暴露在黄麴毒素情形下;影响最钜的是转录过程受抑制,碳水化合物及脂质的代谢以及粒线体呼吸作用的改变。且黄麴毒素与大分子(核酸与蛋白质等)与细胞微器官(粒线体与核醣体)之间又有交互作用,并干扰酵素及其他细胞调节物质(cellular regulators)。 四、黄麴毒素对营养分吸收代谢之影响: 黄麴毒素最主要之标的器官为肝脏;肝脏将之代谢为2.3-oxide,此一代谢具强烈之肝毒性。因肝脏坏死及干扰肝脏正常功能,胆汁产生减少及胰脏酵素分泌减少,致对营养分的吸收不良。此外;本节也将讨论饲料中孟宁素monensin与单宁对黄麴毒素的交互作用。 孟宁素为一种streptomyces cinnamonesis所产生的代谢物,用以控制球 病的发生;鸡只被球 攻击後,日粮中孟宁素与黄麴毒素同时存在所导致的盲肠病变(cecal lesion)异於单纯球 感染者,造成田间诊断鸡只病变上的困扰。日粮中单宁含量一般之许可限量为2%以下,但此一浓度下之单宁与黄麴毒素存在具加成性的毒性产生。 (一)黄麴毒素与单宁、monensin的交互作用 Dale et al. (1980)连续使用两个试验来探讨单宁与黄麴毒素的关系。试验一使用试 级单宁来源,试验二则以BR-64品种之高梁来供应单宁,结果如日粮中含0.5%及1.0%单宁和2.5ppm黄麴毒素时;体重显着下降,饲料换肉率(FCR)亦变差,显示单宁与黄麴毒素有加成性毒害作用。 Tamir and Alumot (1970)指出,单宁与蛋白质在胃肠内形成不溶物,且Osborne et al. (1976)发现黄麴毒素造成胰脏之解脂鶤,淀粉鶤,胰蛋白鶤活性下降,推断可能由此两种原因加成所致。 Wyatt et al. (1975)探讨日粮中之黄麴毒素及孟宁素时,再以球 (E. tenella)卵囊接种後;计算6日内之鸡只死亡率,日粮含孟宁素组死亡率为10%;当添加黄麴毒素时提高至30%;此时若无孟宁素添加则死亡率高达60%。故黄麴毒素与球 症具有协同增加鸡只死亡率之作用。此外,病理学判定上,两者同时存在时其病变记分(lesion score)较单独以球 感染之病变记分为低,病变记分是以盲肠中凝血块及盲肠扩张程度而定。Doerr et al. (1974)指出aflatoxin干扰血凝,特别是外源性和一般血凝途径,可能因此造成凝血慢且盲肠较不呈扩张的外观如图4。(A)为只感染E. tenella可见盲肠凝血及扩张;(B)为E. tenella+2.5ppm黄麴毒可见盲肠内凝血少且扩张小。此一特殊病变,在田间常被误判为非球 感染引起之死亡。 (二)aflatoxin对胆沈钙化固醇、碳水化物代谢及铁吸收的影响 Bird et al. (1978)指出,aflatoxin含阻碍VitD3在体内之代谢;是因肝脏破坏後;产生之25-hydroxylase减少所致,同时胫骨中灰分量减少,资料经复回归方程式(multiple regression equation)导出一公式: Y=40.44962-0.27464X1+0.03197X2 X1:为AFB1的ppm X2:为VitD3 ICU 图5为回归平面图,图中可知黄麴毒素提高,骨灰分减少,VitD3增加,骨灰分增加。由方程式计算知日粮中含1ppm AFB1时,须额外增加8.84 ICU VitD3方可使骨中灰分维持不变。 |
| 体内之碳水化合物代谢亦变黄麴毒素影响。Raj et al (1970)的研究发现;黄麴毒素会造成鸡只肝醣生成量减少,肝脏无法迅速将葡萄糖转为肝醣,故其葡萄糖耐性减低。另外在碳水化合物代谢过程中相当重要的醣解作用(glycolysis),醣解作用主在鸡肝脏及肌肉中进行以产生高能化合物(ATP)供能量所需。黄麴毒素降低醣解过程中几个重要之速率决定酵素(key enzyme)如hexokinase,phosphoglucoisomerase,fructosediphosphate aldolase,pyruvate kinase等醣素活性;如表2所示。可见黄麴毒素症爆发时;鸡只无法藉由醣解来产生能量;但糖质新生(Gluconeogenesis)过程中重要之key enzyme,pyruvate carboxylase及PEP carboxykinase活性显着地升高。故鸡只对能量来源转而由胺基酸代谢供应,如表3所示。 |
| 表二 黄 麴 毒 素 B1 对 鸡 肝 中 某 些 醣 解 酵 素 的 影 响 | |||
| Enzyme activity | Control | Treated | P value |
| Hexokinase
Units/g liver Specific activity Phosphoglucoisomerase Units/g liver Specific activity Fructosediphosphate aldolase Units/g liver Specific activity Pyruvcate kinase Units/g liver Specific activity Lactate dehydrogenase Units/g liver Units/g protein |
2.30±0.16 19.22±1.41
145.90±4.77 810.0±20.30
5.97±0.19 52.33±1.75
6.88±1.33 2.67±0.40
188.4±20.0 141.7±31.0 |
0.56±0.04 6.09±0.52
69.40±1.53 680.0±19.5
3.27±0.05 34.77±1.77
61.00±2.16 24.20±0.82
1.570±2.00 0.540±190 |
0.001 0.01
0.05 0.05
0.001 0.001
0.001 0.001
N.S N.SU |
| 比活性以units/g蛋白质表示,每一数值是6只鸡之平均±标准机率差,P≤0.05表示显着差异。N.S.代表无显着差异。 (Raj et al., 1974) | |||
|
表三 黄 麴 毒 素 B1 鸡 肝 中 fructose-1,6-diphosphatase , pyruvate carboxylase 和 phosphoenol pyruvate carboxykinase 之 影 响 |
|||
| Enzume assayedy | Control | Treated | P value |
| Fructosediphosphatase
Units/g liver Specific activity Pyruvate carboxylase Units/g liver Specific actuvity PEP carboxykinase Units/g liver Specific activity |
11.15±0.46 92.42±3.18
275.0±56.83 5,220±240
2.4×105 21×105 |
12.35±2.50 93.31±3.61
688.0±21.67 31,730±1,380
4.4×105 47×105 |
N.S. N.S.
0.001 0.001
0.001 0.001 |
| Specific activity is experssed as units/g protein. Each value is the mean±S.E. for 6 birds. P≤0.05 is considered significant. N.S. means not significant. (Raj et al., 1974) | |||
|
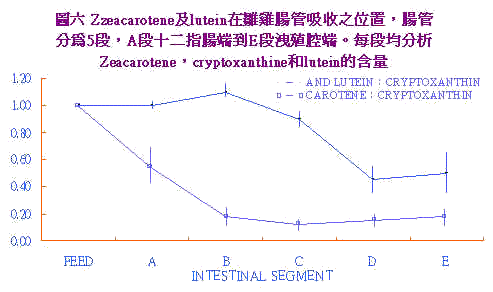
黄麴毒素中毒所造成之贫血,先前之研究(ForgacsCarel, 1962)认为是形成不全性贫血(aplastic)。Tung et al. (1975)及Smith et al. (1970)均指出随aflatoxin剂量之提高血球容积比(PCV)随之下降。Tung et al. (1975)认为是溶血性贫血(hemolysis)。Lanza et al (1979)应用同位素59Fe喂与白肉鸡探讨aflatoxin对PCV的影响,结果0~3wks龄黄麴毒素处理组之PCV,显着较低;3~6wks龄时却无此现象出现;此与Lanza et al. (1978)发现随周龄增加;aflatoxin所造成之贫血反应较不敏感。试验中也发现Fe的主动运输减少;主动运输需要transferrin的媒介;transferrin由肝脏所制造;肝脏坏死情形下铁运输即因transferrin之减少而下降,因而有PCV低及贫血现象产生。 五、黄麴毒素引发白鸟症之机制: 黄麴毒素毒性试验中发现会降低血中的胡萝卜素(hypocarotenoidmia),引起白鸟症(pale bird syndrome)。白鸟症之鸡只可视粘膜,如眼睑、冠、瞬膜、皮肤、脚胫之色素褪除(depletiom);显现出苍白外观。然除黄麴毒素会引起外,其他如赭黄毒素(Ochratoxin)、球 感染、里奥病毒(Reovirus)和日粮不平衡均会发生。本节乃讨论黄麴毒素诱发白鸟症之机转。 Osborne et al. (1982)以各种霉菌毒素(T2-toxin, aflatoxin, Ochratoxin)喂予白肉鸡发现有低胡萝卜血症;因此认为aflatoxin可能导致白鸟症之发生。因对carotenoids的吸收及沈积机构了解甚少,一般咸认为其伴随脂肪被消化吸收及代谢。Tyczkowski et al. (1986a)试验指出鸡对玉米黄素(cryptoxamthin)吸收差,以其为参考值,比较zeacarotene/cryptoxanthine及lutein/cryptoxanthine之比。在各段肠道消失之位置。结果得知zeacarotene和叶黄素(lutein)吸收部位不同,前者主在十二指肠及空肠,後者则为回肠中後段,如图6。 |
|
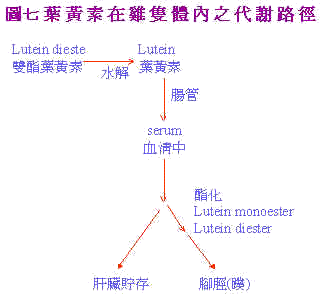
Tyczkowski et al. (1986b)在探讨叶黄素於体内运输,代谢及沈积型式时,指出lutein diester进入鸡体内後,於肠中进行去醯基化(deacylation)後以lutein及单酯(lutein monoester)形式进入血中,肝脏所堆积之型式为游离叶黄素(free lutein)占88%,脚蹼部份主要为lutein diesten及monoester。如图7及表4所示。
|
| 表五 各 种 型 态 叶 黄 素 在 体 内 之 分 布 情 形 | |||
| 部 份 | Lutein diester | Lutein monoester | Free lutein |
| 空肠黏膜
血 清 肝 脏 脚 蹼 |
少量
少量 6% 66% |
6%
1% 6% 8% |
94%
99% 88% 26% |
|
Tyczkowski et al. (1987)证实aflatoxin使得lutein diester吸收大为减少,且影响各部份中lutein之型式,遂有白鸟症之产生。Schaeffer et al. (1988a)即利用 物动力学方法(pharmacokinetic method)探究表皮中lutein酯化沉积的速率。以K1,K2,K3,代表速率常数: 血清中游离叶黄素→K1→表皮 中游离态叶黄素→ K2→表皮中单酯叶黄素→K3→表皮中双酯叶黄素 结果显示经黄麴毒素处理後K2,K3值降低,显示表皮之醯化(acylatien)对黄麴毒素相当敏感,但是K1不被影响。由於肝脏中之主要为游离态之叶黄素;但黄麴毒素致使肝脏中主要为酯化型态使待运送到表皮的Lutein无法进行。如表5以上所知均为对Lutein蓄积的抑制;是否黄麴毒素亦会加速色素在体内之褪除;Schaeffer et al. (1988b)研究显示黄麴毒素对空肠中、血清中及脚蹼中色素褪除均无影响。 物动力学研究亦显示,黄麴毒素对K1,K2及K3值之模式型态(K1最慢,K2次之,K3最慢)与对照组相同。如表6所示。 |
| 表五 黄麴毒素对表皮色素沈积反应之速率常数的影响 | ||
| Rate速率A | Control | Aflatoxin B |
| ──────── (day -1) ──────── | ||
| K1
K2 K3 |
2.51×10-1±0.03a
2.65×10-1±0.01a 0.83×10-1±0.01a |
2.08±10-1±0.03a
0.05×10-1±0.01b 2.15×10-1±0.01b |
| a,b同一行中有不同上标者表显着差异(p<0.05)
A 速率常数的代号:K1=Lutein的形成;K2=lutein monoester的形成K3=lutein diester的形成 B 日粮中含2ppm之黄麴毒素 (Schaeffer et al., 1988a) |
||
| 表六 黄麴毒素对表皮色素褪除反应之速率常数的影响 | ||
| Constant | Control | Aflatoxin AB |
| ────── (day -1) ────── | ||
|
K1(Diester→monoester) K2(Monoester→lutein) K3(Lutrin→serum lutein) |
7.4×10-2±0.006a
3.6×10-2±0.15b 2.4×10-1±0.01c |
6.3±10-2±0.008a
4.2×10-2±0.1a 2.3×10-1±0.01b |
| a-b同一中有不同上标者表显着差异(P<0.05)
A 日粮中含4ppm之黄麴毒素 B 对照组之各别速率常数与相关之黄麴毒素处理组速率常数比较,显示无显着差异。 (Schaeffer et al., 1988a) |
||
|
Day et al. (1958)指出,日粮中添加5%牛油可增加色素沈积,Health et al. (1972)於日粮 加大豆油时可刺激oxycarotenoid在背部皮肤之蓄积。 Han et al. (1987)喂予玉米变异种,含高量油可刺激脚胫和血浆色素沈着。 Schaeffer et al (1990)指出,日粮中脂肪含量增加,血清中lutein及lutein代谢物3-oxolutein均增加且於6%脂肪添加时达到最高。如表7所示日粮中含有aflatoxin时亦有相同之结果。显示脂肪增加可促进lutein溶於脂肪及乳糜微粒(micelle)之形成。 |
| 表七 日粮添加各种水准脂肪对黄麴毒素症鸡只之血清oxycarotenoids的影响 | ||||
| Fat dietA % | 3-OxoluteinB | LuteinB | ||
| - | + | - | + | |
| 0
2 4 6 8 10 |
0.36±0.06d
1.09±0.03c 1.56±0.05b 1.84±0.07a 1.77±0.06a 1.75±0.01a |
0.08±0.01b
0.28±0.05b 0.46±0.08b 0.91±0.16a 0.92±0.13a 1.00±0.19a |
5.5±4e
17.1±0.4d 24.1±.1.2c 32.0±.1.2a 28.6±0.6ab 27.1±.1.4bc |
0.7±0.1c
3.0±0.3b 5.7±0.12b 11.1±.1.8a 10.8±.1.2a 14.3±.4a |
| a-b
同一列中有不有同上标者表显着差异(P<0.05)。
A 鸡只喂予的白玉米黄豆粉和淀粉再水准之棉子油的基础日粮3周後。一组中10只鸡平均浓度+SE的值。 B +或-代表有或无1.4ppm黄麴毒素之添加。 (Schaeffer et al., 1990) |
||||
|
脂肪来源中富含饱和短链脂酸(如capric acid C10:0;lauric acid C12:0)对於因黄麴毒素而造成lutein吸收下降具有拮抗效果;而长链不饱和脂酸效果较饱和短链者为差。黄麴毒素因降低鸡只对脂肪消化过程中形成乳糜微粒之能力,是以micelles形成减少。故有Osborne和Hamilton (1981)所观察到之脂肪痢现象产生。 脂肪水准高时,lutein吸收增加,反应出oxycartenoids吸收须脂肪来溶解,并形成micelles再将oxycarotonoids运入肠细胞。 黄麴毒素剧烈的降低血清中脂质含量(Fung et al., 1970)故亦可能降低内源性脂肪和单酸甘油酯在胆汁中形成的量。在日粮脂肪缺乏时,内源性的疏水性及亲水性脂质之形成相形减少,仅能提供有限度的micelles形成及有限的日粮oxycarotenoids的运输。 而日粮中提高脂肪对黄麴毒素的毒害有抵抗的效果,乃增加本来就有限的疏水性及亲水性脂质(受黄麴毒素而减少者);且短链饱和脂酸可作为形成micelle中心之疏水端,较佳於其他种类脂酸。 六、结 论 1.黄麴毒素为Aspergillu flavus与Aspergillus parasiticus利用醋酸盐作为基质,经Polyketide生合成代谢路径,所合成之次级代谢产物;在合成过程中锌离子与产毒素量具有密切之关系;可能为其中间代谢酵素之辅因子;黄麴毒素较重要的有B1、B2、G1、G2;其中以黄麴毒素B1之毒性最强;危害亦最大。 2.黄麴毒素进入物体内有两种不同之代谢径路。分别为发生於啮齿类动物及哺乳类动物之肝脏;黄麴毒素B1经代谢为毒性极强之黄麴毒素epoxide再与核酸和蛋白质结合,并干扰DNA的转录、转译过程。故具可导致突变、致畸胎与致癌性。 3.黄麴毒素造成肝坏死、肝细胞退化、胆小管上皮细胞增生,胆汁产生减少及胰脏酵素分泌变少,故影响到动物体对营养分之吸收和代谢。如阻碍维生素D3在体内之代谢;葡萄糖耐性减强;铁吸收减少等。 4.白鸟症的发生乃因黄麴毒素影响叶黄素(lutein)在肠管之运输和醯基化使之与正常之代谢过程不同;使得lutein diester和monoester均沈积於肝脏中无法生去醯基化成free lutein运输至表皮表脚胫中沈积。 但对於色素之褪除;黄麴毒素则无影响。藉由日粮中提供含饱和短链脂酸和不饱和长链脂酸时;可提高叶黄素之吸收;对黄麴毒素之阻碍吸收作用有节约效果(sparing effect)。 七、引用文献 1.杂粮与畜产月刊。12月。1990。 2.世界玉米黄麴毒素研究专辑。1990。台湾区杂粮发展基金会。 3.林茂勇。1991。霉菌毒素学。淑馨出版社。 4.颜国钦。1980。食品安全学。艺轩出版社。 5.Mycotoxins in human and animal health;Aflotoxin p8-98. 1977. Pathotox publisher. inc. 6.Aso, T. et al. 1970. J. Am. Chem. Soc., 85:1706-1707. 7.Bird, F. S. et al. 1978. Poult. Sci., 57:1293-1296. 8.Campbell, M. L. et al. 1983. Poult. Sci., 62:2138-2144. 9.Chang. C. F. et al. 1979. Poult. Sci., 58:562-566. 10.Dalle, N. M. et al. 1980. Poult Sci 69:2417-2420. 11.Day, E. J. et al. 1958. Poult. Sci., 37:1373-1381. 12.Doerr. J. A. et al. 1974. Poult. Sci., 53:1728-1734. 13.Forgacs, J. et al. 1962. Adv. Vet. Sci., 7:273-382. 14.Gupta, S. K. et al. 1976. Appl. Enviro. Microl., 32:324. 15.Gupta and Venkitabramanian. 1977. Microbiol. 18:27-34. 16.Han, Y. C. M. et al. 1987. poult. Sic., 66:103-111. 17.Heath, J. J. et al. 1972. Poult. Sci., 51:502-506. 18.Huff, W. E. et al. 1986. Poult Sci., 65:1891-1899. 19.Lanza, G. M. et al. 1978. Poult. Sci., 59:282-288. 20.Lanza, G. M. et al. 1979. Poult. Sci., 59:1439-1444. 21.Lee, L. S. et al. 1974. J. Assoc. Off, Anal., 57:626-631. 22.Maggon, K. K. et al. 1977. Bacteriol. Rev., 41:822-855. 23.Osborne, D. J. et al. 1976. Poult. Sci., 54:1802. 24.Osborne and Hamilton. 1981. Poult. Sci., 60:1398-1402. 25.Osborne and Hamilton. 1982. Poult. Sci., 61:1646-1652. 27.Raj, H. G. et al. 1974. Environ. Phisiol. Biochem., 4:181-187. 28.Schaeffer, J. L. et al. 1988. Poult. Sci., 67:1080-1088. 29.Schaeffer, J. L. et al. 1988. Poult. Sci., 67:619-625. 30.Schaeffer, J. L. et al. 1990. Poult. Sci., 69:53-59. 31.Smith, J. W. et al. 1970. Poult. Sci., 49:207-215. 32.Swenson, D. H. et al. 1973. Biochem. Biophys. Res. Commun., 53:1260-1267. 33.Swenson, D. H. et al. 1973. Biochem. Biophys. Res. Commun., 60:1036-1043. 34.Tamir, M. et al. 1970. J. Nutr., 100:573-580. 35.Tung, H. T. et al. 1972. Appl. Pharmacol., 22:27-104. 36.Tung, H. T. et al. 1975. Poult. Sci., 54:1962-1969. 37.Tyckowski, J. K. et al. 1991. Poult. Sci., 70:2275-2279. 38.Tyckowski, J. K. et al. 1986a. Poult. Sci., 65:1526-1531. 39.Tyckowski, J. K. et al. 1986b. Poult. Sci., 66:1184-1188. 40.Wyatt, R. D. et al. 1975. Poult. Sci., 54:1065-1070. |